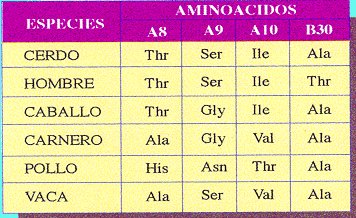
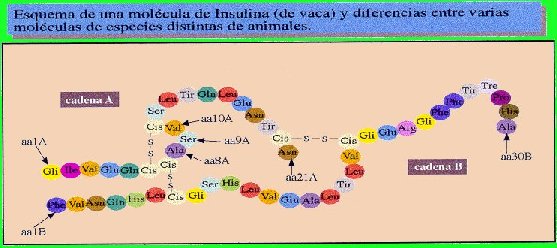
Sin embargo, pudo observar que cada tanto aparecía alguna variante que no estaba presente ni en los progenitores ni en ningún antecesor de estas mismas plantas.
A partir de lo ocurrido De Vries planteo una hipótesis, donde determinó que estas variantes surgían como resultado de cambios súbitos en los genes y que la variante producida por un gen cambiado se transmitía luego a la progenie, como lo hace cualquier otra característica hereditaria. De Vries denominó mutaciones a estos cambios hereditarios repentinos, y a los organismos que exhibían estos cambios, mutantes. Los conceptos propuestos por de Vries no resultaron tan erróneos, el concepto de mutación como fuente de la variación genética demostró ser de suma importancia, aunque la mayoría de sus ejemplos no eran muy válidos.
Hoy se sabe que las mutaciones son cambios abruptos en el material genético. Como resultado de las mutaciones, existe una amplia gama de variabilidad en las poblaciones naturales. En un ambiente heterogéneo o cambiante, una variación determinada puede darle a un individuo o a su progenie una ligera ventaja, como así también un efecto negativo provocando la extinción de una especie. En consecuencia, aunque las mutaciones no determinan la dirección del cambio evolutivo, constituyen la fuente primaria y constante de las variaciones hereditarias que hacen posible la evolución.
Para resumir entonces:
"Una mutación es un cambio heredable en el material genético de una célula".
En la naturaleza las mutaciones se originan al azar y, aunque las causas siguen siendo inciertas, se conocen bastantes agentes externos, mutágenos, que pueden producir mutaciones como: las radiaciones ambientales y sustancias químicas. Una mutación en una célula somática, puede provocar alteraciones en el organismo en el que se presente; pero desaparece en el momento en que muere el individuo en que se originó. Sin embargo, las mutaciones en las células sexuales, óvulos y espermatozoides, pueden transmitirse como rasgos hereditarios diferenciadores a los descendientes del organismo en los que tuvo lugar la mutación.
Se distinguen varios tipos de mutaciones en función de los cambios que sufre el material genético.
1. Mutaciones cromosómicas. Este tipo de mutaciones provoca cambios en la estructura de los cromosomas.
- Deleción. Implica la pérdida de un trozo de cromosoma; los efectos que se producen en el fenotipo están en función de los genes que se pierden.

- Duplicación. En este caso existe un trozo de cromosoma repetido.

2. Mutaciones genómicas. Este tipo de mutaciones afecta a la dotación cromosómica de un individuo, es decir, los individuos que las presentan tienen en sus células un número distinto de cromosomas al que es propio de su especie. No son mutaciones propiamente dichas, porque no hay cambio de material genético, sino una aberración, la cual suele ser el resultado de una separación anormal de los cromosomas durante la meiosis, con lo que podemos encontrarnos individuos triploides (3n), tetraploides (4n), etc.
Estos poliploides así formados son genéticamente muy interesantes en las plantas cultivadas, y hoy en día la mayoría de variedades gigantes de fresones, tomates, trigo, ... que existen en el mercado, tienen este origen.
En el hombre, existen varios síndromes provocados por la falta de separación de una pareja de cromosoma homólogos durante la meiosis, con lo cual permanecen unidos y se desplazan juntos a un mismo gameto provocando lo que se denomina trisomía, es decir un individuo con un cromosoma triplicado.
En los siguientes esquemas, tenemos las trisomías más frecuentes tanto en los autosomas, como en los cromosomas sexuales.
Síndrome
|
Tipo de mutación
|
Características y síntomas de la
mutación
|
Síndrome de Down
|
Trisomía 21 |
Retraso mental, ojos oblicuos, piel rugosa, crecimiento retardado
|
Síndrome de Edwars
|
Trisomía 18 |
Anomalías en la forma de la cabeza, boca pequeña, mentón huido,
lesiones cardíacas.
|
Síndrome de Patau
|
Trisomía 13 o 15
|
Labio leporino, lesiones cardíacas polidactilia.
|
Síndrome de Klinefelter
|
44 autosomas + XXY
|
Escaso desarrollo de las gónadas,
aspecto eunocoide.
|
Síndrome del duplo Y
|
44 autosomas + XYY
|
Elevada estatura, personalidad infantil, bajo coeficiente
intelectual, tendencia a la agresividad y al comportamiento antisocial.
|
Síndrome de Turner
|
44 autosomas + X
|
Aspecto hombruno, atrofia de ovarios, enanismo.
|
Síndrome de Triple X
|
44 autosomas + XXX
|
Infantilismo y escaso desarrollo de las mamas y los genitales
externos.
|
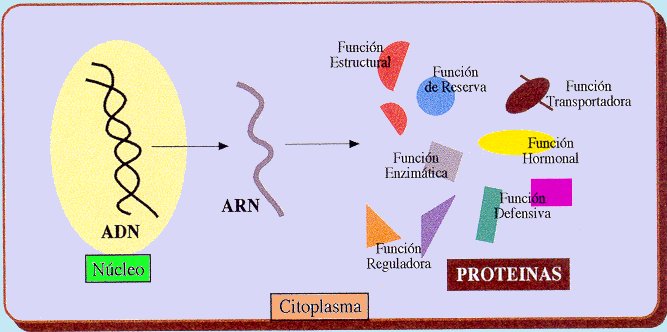
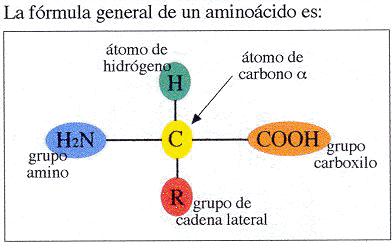
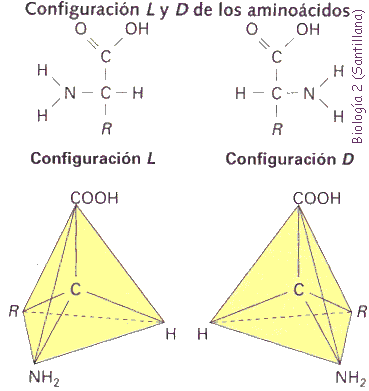
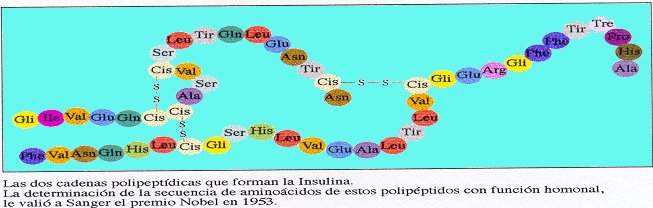
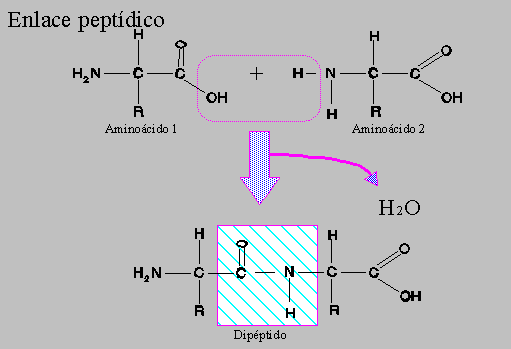
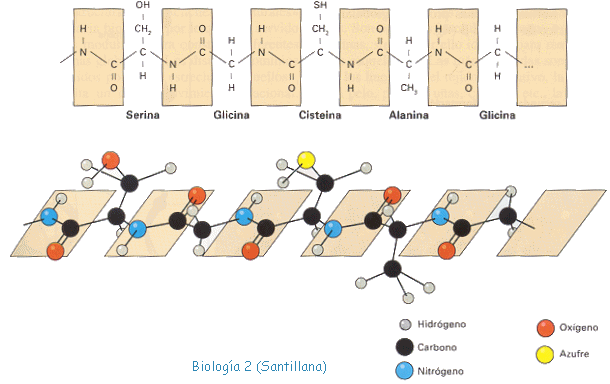
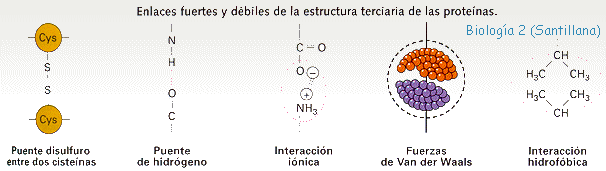
3. Mutaciones génicas. Son las verdaderas mutaciones, porque se produce un cambio en la estructura del ADN. A pesar de todos los sistemas destinados a prevenir y corregir los posibles errores, de vez en cuando se produce alguno en la réplica, bien por colocarse una Citosina (C) en lugar de una Timina (T), o una Adenina (A) en lugar de una Guanina (G); o bien porque el mecanismo de replicación se salta algunas bases y aparece una "mella" en la copia. O se unen dos bases de Timina, formando un dímero.


Aunque se trate de un cambio de un nucleótido por otro, supondrá una alteración en la secuencia de un gen, que se traduce posteriormente en una modificación de la secuencia de aminoácidos de una proteína.
Al transcribirse la mutación, al menos un triplete del ARNm , se encuentra modificado y su traducción da lugar a que se incorpore un aminoácido distinto del normal en la cadena polipeptídica. Es un cambio que aunque la mayoría de las veces va a ser perjudicial, en contadas ocasiones puede provocar que mejore un gen y gracias a esta característica se sintetice una proteína distinta, que tenga propiedades distintas o participe en la formación de estructuras más eficaces.
En estos casos raros, pero esenciales para la evolución de las especies, los individuos portadores de la mutación poseen ventajas adaptativas respecto a sus congéneres, por lo que el gen mutado es posible que con el tiempo, y gracias a la selección natural, sustituya al gen original en la mayoría de los individuos que componen la población.
Por el Ing. Agr. Carlos González